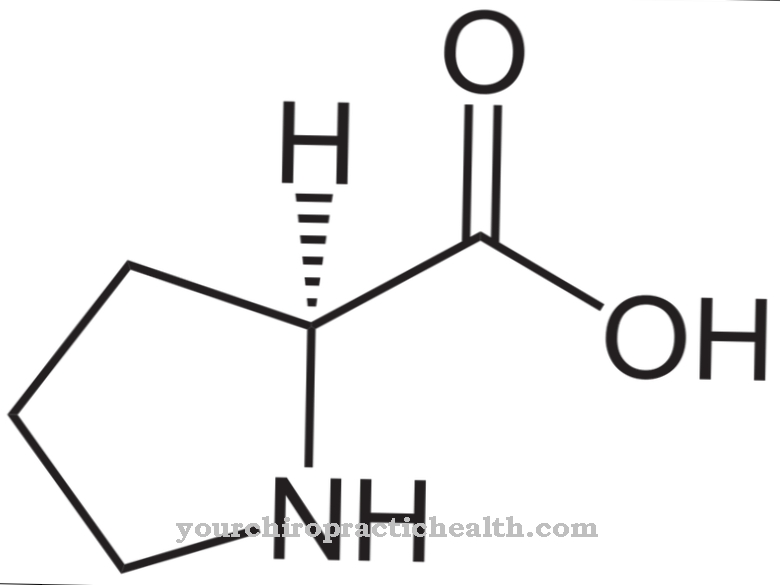
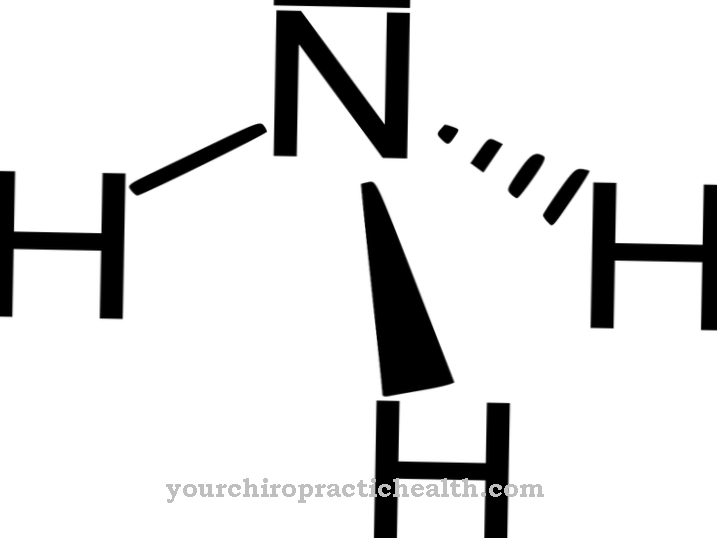
Цитидиновата принадлежи към нуклеозидите и е изграден от цитозин на нуклеиновата основа и захарната рибоза. Образува основна двойка с гуанозин чрез водородни връзки. Той също играе централна роля в метаболизма на пиримидин.
Какво е цитидин?
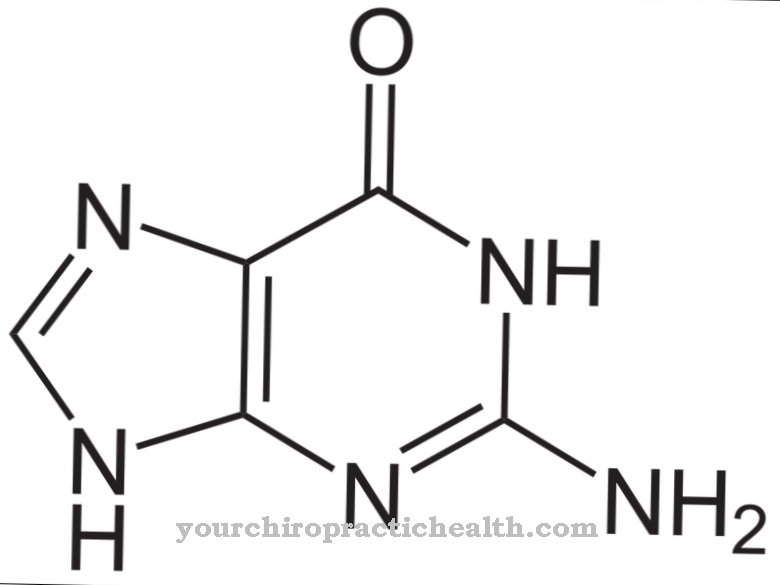
Цитидинът е нуклеозид, който се състои от цитозин и рибоза. Освен аденин, гуанин и тимин, цитозинът на азотна основа участва в синтеза на нуклеинови киселини. Фосфорилирането на цитидин произвежда цитидин монофосфат (CMP), цитидин дифосфат (CDP) или цитидин трифосфат (CTP).
Цитидин монофосфатът е нуклеотид в РНК. Две пуринови и две пиримидинови основи участват в структурата на нуклеиновите киселини, като тиминът се обменя на урацил в РНК. Аденинът и гуанинът принадлежат към пуриновите основи, докато тиминът, цитозинът и урацилът принадлежат към основите на пиримидина. Цитидин дезаминазата може да деаминира цитидин до уридин. Уридинът е нуклеозид, произведен от рибоза и урацил. Той може също да бъде фосфорилиран до уридин монофосфат.
Уридин монофосфатът също е важен нуклеотид за РНК. Освен това CDP и CTP също активират групи за синтеза на лецитин, цефалин и кардиолипин. Чистият цитидин присъства като водоразтворимо твърдо вещество, което се разлага при 201 до 220 градуса. Тя може да бъде каталитично разградена до цитозин и рибоза чрез ензима пиримидин нуклеозидаза.
Функция, ефект и задачи
Цитидинът играе централна роля в метаболизма на пиримидин. Пиримидинът осигурява основната структура на пиримидиновите основи цитозин, тимин и урацил, които се срещат в нуклеиновите киселини. Тиминът в РНК се заменя с урацил.
Урацил се получава също чрез дезаминиране на цитидин с цитидин дезаминаза. Химическите преобразувания между трите основи на пиримидин са от централно значение за възстановителните процеси в ДНК и епигенетичните промени. В контекста на епигенетиката различни свойства се променят от влиянието на околната среда. Генетичният материал обаче не се променя. Модификационните промени в организма се причиняват от различната експресия на гени. Процесите на диференциация на телесните клетки за образуването на различни клетъчни линии и органи също представляват епигенетичен процес, В зависимост от типа на клетките се активират или деактивират различни гени.
Това става чрез метилиране на цитидиновите бази в ДНК. По време на метилирането се образува метилцитозин, който чрез дезаминиране може да се превърне в тимин. Комплементарният нуклеобазен гуанин в противоположната двойна верига позволява грешката да бъде разпозната и тиминът да бъде заменен отново с цитозин. Гуанинът обаче може да бъде заменен и за аденин, което води до точкова мутация. Ако неметилираният цитозин се дезаминира, се получава урацил. Тъй като урацил не се появява в ДНК, той веднага се замества с цитозин. Вместо цитозин скоростта на мутация поради метилиране е леко повишена.
В същото време все повече и повече гени се изключват чрез метилиране, така че клетките в клетъчната линия да станат по-специализирани. В процесите на ремонт възстановителните ензими се основават на оригиналната верига на ДНК, която те разпознават чрез по-висока степен на метилиране. Допълващата нишка се изгражда и въз основа на информацията, съхранявана там. Грешките при инсталиране се коригират незабавно. Освен това ензимът AID (активирана индуцирана цитидин дезаминаза) много специфично катализира дезаминирането на цитидиновите групи до уридиновите групи в едноверижната ДНК. Появяват се соматични хипермутации, които променят последователността на антителата на В клетки. Тогава се избират съвпадащите В клетки. Това дава възможност за гъвкав имунен отговор.
Образование, възникване, свойства и оптимални стойности
Цитидинът е междинен продукт на метаболизма на пиримидин. Като изолирана връзка няма значение. Както вече споменахме, той е съставен от цитозин на нуклеиновата основа и рибозата на захарната четворка. Организмът може сам да синтезира цитозин.
Въпреки това, неговият синтез е много енергоемък, така че той се възстановява от градивни елементи на нуклеиновата киселина в рамките на пътя на спасяване и може да бъде реинтегриран в нуклеинови киселини. Когато основата се разгради напълно, се получават въглероден диоксид, вода и урея. Той присъства като нуклеозид в РНК. В ДНК цитозинът се свързва с дезоксирибоза, така че нуклеозидният дезоксицитидин се среща тук като градивен елемент.
Болести и разстройства
Метилиранията на цитидиновите остатъци от ДНК са много важни за маркирането, за да се разделят различни биохимични процеси. Въпреки това могат да възникнат грешки и при метилиране, които водят до заболяване.
В случай на дефектни метилирания могат да се задействат както повишени, така и намалени генни активности, които не отговарят на изискванията. Тези модели на метилиране се предават по време на клетъчното делене. В дългосрочен план настъпват промени, които могат да доведат до заболявания. Например, някои туморни клетки имат различни структури на метилиране, които не се срещат в здрави клетки. Например, метилирането може да блокира определени гени, които кодират ензими, регулиращи растежа. Ако тези ензими липсват, може да настъпи инхибиран растеж на клетките. Това се отнася и за ензимите, които инициират подредена клетъчна смърт (апоптоза), когато възникнат клетъчни дефекти.
Насочено въздействие на метилирането на ДНК все още не е възможно. Съществуват обаче проучвания за пълното деметилиране на туморните клетки, за да се подложат отново на контрола на протеините, регулиращи растежа. Според няколко клинични проучвания е показано, че деметилирането ограничава растежа на тумора при пациенти с остра миелоидна левкемия. Тази процедура е известна още като епигенетична терапия. Процесите на метилиране могат също да играят роля при други заболявания. Поради влиянието на околната среда организмът се адаптира към променените условия с образуването на биологични модификации, основаващи се на метилиране на цитидиновите остатъци от ДНК. Така организмът осъществява процес на обучение, който обаче може да причини и неправилна регулация.



.jpg)


.jpg)









.jpg)